Contact Information
University of Illinois
B107 CLSL
601 S. Goodwin Avenue
Urbana, IL 61801
Research Description
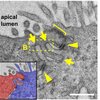
Epithelial Junction
Epithelial cells cover human body cavities and line internal organs, providing protection from biological, chemical, and mechanical insults. In vertebrates, the physiological functions of an epithelium require the assembly of a specialized intercellular structure known as the apical junction.
My laboratory focuses on 3 main topics surrounding the epithelial apical junction. First, we aim to understand the role of the apical junction in providing adhesions between cells. Second, we aim to understand the role of the apical junction in paracellular permeability and ion transport. Third, we aim to dissect the molecular mechanism of junction remodeling. The overall goal of the lab is to understand how the apical junction contributes to epithelial homeotasis and supports the physiological functions of epithelial tissues.
Education
B.A., University of California, Berkeley (Physiology)
Ph.D., University of California, San Francisco (Physiology)
Postdoc, Harvard Medical School, Harvard University
Awards and Honors
Research Career Award, NIDDK, NIH
LAS Impact Award, UIUC
Additional Campus Affiliations
Research Assistant Professor, Cell and Developmental Biology
Research Assistant Professor, Biomedical and Translational Sciences
External Links
Highlighted Publications
Representative Publications
Timothy Morris, Eva Sue, Caleb Geniesse, William M Brieher, Vivian W Tang (2022) Synaptopodin stress fibers and contractomere at the epithelial junction. Journal of Cell Biology Abstract
John X. H. Li, Vivian W. Tang, Kingsley A. Boateng, and William M. Brieher (2021). Actin protrusions push at apical junctions to maintain E-cadherin adhesion. Proceedings of the National Acadamy of Science Abstract ![]()
Vivian W. Tang (2020) Collagen, stiffness, and adhesion: the evolutionary basis of vertebrate mechanobiology. Molecular Biology of the Cell Perspective
Vivian W. Tang, Ambika V. Nadkarni, and William M. Brieher (2020). Catastrophic actin filament bursting by cofilin, Aip1, and coronin. Journal of Biological Chemistry Abstract
John X. H. Li, Vivian W. Tang, and William M. Brieher (2019). Actin protrusions push at apical junctions to maintain E-cadherin adhesion. Proceedings of the National Acadamy of Science Abstract ![]()
Nivetha Kannan and Vivian W. Tang (2018) Myosin-1c promotes E-cadherin tension and force-dependent recruitment of α-actinin to the epithelial cell junction. Highlighted in "May the FORCE be with you" J. Cell. Sci. Abstract ![]()
Vivian W. Tang (2018) Cell–cell adhesion interface: orthogonal and parallel forces from contraction, protrusion, and retraction. F1000Prime Perspective
Vivian W. Tang (2017) Cell-cell adhesion interface: rise of the lateral membrane. F1000Prime Perspective ![]()
Nivetha Kannan and Vivian W. Tang (2015) Synaptopodin couples epithelial contractility to α-actinin-4–dependent junction maturation. J. Cell Biol. 211:407-434. Abstract
Vivian W. Tang and William M. Brieher. (2013) Capping actin barbed-ends by FSGS3/CD2AP stabilizes actin at the adherens junction to strengthen epithelial cohesion and protect permeability barrier. J. Cell Biol. 203:815-833. Abstract
Vivian W. Tang and Willaim M. Brieher. (2012) Alpha-actinin-4/FSGS1 is required for arp2/3-dependent actin assembly at the adherens junction. J. Cell Biol. 196:115-130. Abstract ![]()
Vivian W. Tang (2006) Proteomics and bioinformatic analysis of tight junction reveals an unexpected cluster of synaptic proteins. Biology Direct 1:37. Abstract ![]()
Recent Publications
Morris, T., Sue, E., Geniesse, C., Brieher, W. M., & Tang, V. W. (2022). Synaptopodin stress fiber and contractomere at the epithelial junction. Journal of Cell Biology, 221(5), [e202011162]. https://doi.org/10.1083/jcb.202011162
Li, J. X. H., Tang, V. W., Boateng, K. A., & Brieher, W. M. (2021). Cadherin puncta are interdigitated dynamic actin protrusions necessary for stable cadherin adhesion. Proceedings of the National Academy of Sciences of the United States of America, 118(24), [e2023510118]. https://doi.org/10.1073/pnas.2023510118
Li, J. X. H., Tang, V. W., & Brieher, W. M. (2020). Actin protrusions push at apical junctions to maintain E-cadherin adhesion. Proceedings of the National Academy of Sciences of the United States of America, 117(1), 432-438. https://doi.org/10.1073/pnas.1908654117
Tang, V. W., Nadkarni, A. V., & Brieher, W. M. (2020). Catastrophic actin filament bursting by cofilin, Aip1, and coronin. Journal of Biological Chemistry, 295(38), 13299-13313. https://doi.org/10.1074/jbc.RA120.015018
Tang, V. W. (2020). Collagen, stiffness, and adhesion: The evolutionary basis of vertebrate mechanobiology. Molecular biology of the cell, 31(17), 1823-1834. https://doi.org/10.1091/mbc.E19-12-0709